
W poprzednich dwóch artykułach1 opisałem rewolucję zachodzącą w biologii systemów, w której to dyscyplinie założenia ewolucjonistyczne zastąpiono założeniami opartymi na idei projektu, na przykład założeniem o kluczowej roli teleologii. W niniejszym artykule pokażę, że biologowie coraz częściej odrzucają przekonanie o powszechnym występowaniu marnych projektów w przyrodzie ożywionej. Teraz zazwyczaj przyjmują oni, że struktury i układy biologiczne cechują się wysokim stopniem optymalności.
Oczekiwanie istnienia marnych projektów
Na gruncie logiki standardowego modelu ewolucyjnego przewiduje się, że w biosferze powinno roić się od organizmów mających wadliwe projekty i niefunkcjonalne relikty swojej ewolucyjnej przeszłości. Rację dla takiego przewidywania dobrze przedstawiono w opublikowanym w Wikipedii artykule Argument from Poor Design [Argument z marnego projektu]:
Istnienie „marnego projektu” jest zgodne z przewidywaniami naukowej teorii ewolucji drogą doboru naturalnego. Przewiduje ona, że cechy, które wyewoluowały do pełnienia pewnych funkcji, były następnie ponownie wykorzystywane lub kooptowane do pełnienia innych funkcji albo ulegały całkowitej eliminacji. Ten suboptymalny stan ma związek z niezdolnością mechanizmu dziedziczenia do eliminowania poszczególnych reliktów procesu ewolucji.
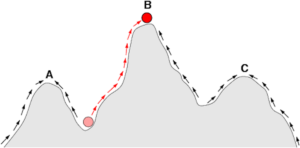
Ujmijmy to w kategoriach krajobrazu przystosowania: dobór naturalny zawsze będzie parł „w górę”, ale gatunek zwykle nie może przejść od niższego szczytu do wyższego, jeśli najpierw nie przejdzie przez dolinę2.
{kind=link}
Oczekiwanie istnienia marnych projektów nie jest po prostu subiektywnym, opartym na intuicji wnioskiem, lecz w ścisły sposób wykazano jego słuszność za pomocą modeli obliczeniowych. Jeden z takich modeli, opracowany przez Davida W. Snoke’a, Jeffreya Coxa i Donalda Petchera3, pokazał, dlaczego procesy ewolucyjne, które umożliwiają wzrost złożoności, muszą generować duże ilości śmieciowego DNA i niefunkcjonalnych elementów. Szczegóły ich modelu są złożone, ale leżąca u jego podstaw logika jest prosta.
Aby mogły pojawić się złożone innowacje, w organizmach musi powstawać niefunkcjonalny DNA i musi on trwać w populacji, aż w końcu pojawią się funkcjonalne sekwencje. Takie dodatki do genomu mogą tworzyć się wskutek duplikacji i dalszego wielokrotnego mutowania genów. Śmieciowy DNA nieuchronnie będzie się kumulował i obejmie znaczną część genomu. To dlatego biologowie wcześniej zakładali, że śmieciowy DNA składa się nawet na 97 procent genomu człowieka4.
Również powstawanie złożonych struktur (na przykład maszyn molekularnych) wymaga tworzenia niezliczonych, sprawdzanych metodą prób i błędów układów cząsteczek lub tkanek, aż pojawi się coś przynoszącego korzyści. Większość prób będzie przynosiła skutki niefunkcjonalne albo niewydajne. W związku z tym jedynie mniejszość struktur i układów biologicznych powinna cechować się wysokim stopniem optymalności.
Główny argument przeciwko teorii inteligentnego projektu
Najwyraźniejsza różnica w przewidywaniach teorii inteligentnego projektu i teorii niekierowanej ewolucji dotyczy istniejącej w przyrodzie ożywionej proporcji suboptymalnych lub niefunkcjonalnych projektów do projektów optymalnych. Filozof Philip Kitcher kładł na to nacisk w książce Living with Darwin: Evolution, Design, and the Future of Faith [Życie z Darwinem. Ewolucja, projekt i przyszłość wiary]. Podawał on przykłady – w jego przekonaniu – nieudolnych, niekompetentnych projektów, uznając je za główny argument za odrzuceniem teorii inteligentnego projektu:
Gdybyś był utalentowanym inżynierem mogącym zaprojektować wieloryba od zera, to prawdopodobnie nie wyposażyłbyś go w szczątkową miednicę. […] Gdybyś projektował ludzkie ciało, to z pewnością mógłbyś udoskonalić kolano. A gdybyś projektował genomy organizmów, to bez wątpienia nie wypełniłbyś ich śmieciami5.
Podobnie argumentuje biolog Nathan H. Lents w książce Człowiek i błędy ewolucji. Niepotrzebne kości, zepsute geny i inne niedoskonałości ludzkiego ciała. Jego zdaniem „nieudolne” projekty widoczne w całym ciele człowieka wskazują, że nie jesteśmy wytworem inteligentnego projektu, lecz niekierowanego procesu ewolucji:
Do trzeciej kategorii należą te ludzkie defekty, które można przypisać jedynie ograniczeniom samej ewolucji. Każdy gatunek może ulepszać ciało, jakim dysponuje, wyłącznie w drodze bardzo drobnych zmian, które występują przypadkowo i rzadko. Odziedziczyliśmy potwornie nieefektywne struktury, których nie da się zmienić. Z tego właśnie powodu przez ciasną przestrzeń naszego gardła przechodzi zarówno pożywienie, jak i powietrze. Z tego samego powodu w stawach skokowych mamy siedem rozrzuconych bez ładu i składu niepotrzebnych kości. Naprawienie któregokolwiek z tych błędów konstrukcyjnych wymagałoby znacznie poważniejszej interwencji niż pojedyncze mutacje. [Przypuszczenie, że te organizmy żywe zostały oddzielnie stworzone, jest równoznaczne z uznaniem, że tworzący je byt jest kapryśnym, nieudolnym, miernym inżynierem, czyli – inaczej mówiąc – nieinteligentnym projektantem]6.
Zmieniające się perspektywy
Większość podawanych przez Kitchera, Lentsa i innych sceptyków przykładów rzekomo marnego projektu zostało jednak podważonych7. Pozostałe przypadki zwykle dotyczą degradacji niegdyś optymalnych projektów albo stanowią przykłady błędu polegającego na umieszczaniu niedoskonałości w lukach wiedzy8.
Domniemane przykłady marnych projektów to zazwyczaj wyraz opinii kanapowych krytyków, którzy w nikłej mierze rozumieją ustalenia zawarte w literaturze fachowej i nie mają kompetencji w zakresie inżynierii. Na przykład – co bezpośrednio przeczy twierdzeniom Kitchera i Lentsa – inżynierowie powszechnie wielokrotnie wykorzystują motywy projektowe na różne sposoby i okazuje się, że takie ponowne wykorzystanie motywu projektowego ma miejsce również w przypadku miednicy wieloryba9. Specjaliści medyczni oraz inżynierowie wykazali ponadto, że ludzkie kolano oraz staw skokowy są optymalnie i znakomicie zaprojektowane10. W tych strukturach inżynierowie szukali nawet inspiracji do projektowania sztucznych kończyn11.
Co więcej, dzięki ustaleniom projektu ENCODE wiemy obecnie, że większa część ludzkiego genomu jest funkcjonalna12. Niszczycielskie następstwa tego odkrycia dla teorii ewolucji nie umknęły oczywiście uwadze ewolucjonistów. Biochemik Dan Graur otwarcie stwierdził:
Jeśli w genomie człowieka nie ma śmieciowego DNA, na co wskazują wyniki uzyskane w ramach projektu ENCODE, to długi, niekierowany proces ewolucji nie może wyjaśnić ludzkiego genomu. Jeśli zaś organizmy są zaprojektowane, to należy oczekiwać, że cały DNA – albo możliwie największa jego część – jest funkcjonalny. Jeśli wyniki uzyskane w ramach projektu ENCODE są słuszne, to teoria ewolucji jest błędna13.
„Podstawowe zasady optymalności”
Co równie ważne, biologowie systemów wiedzą obecnie, że założenie optymalności projektu prowadzi do najpłodniejszych badań. Na przykład Nikolaos Tsiantis, Eva Balsa-Canto i Julio R. Banga opracowali model przeznaczony do badania układów biologicznych, którego podstawą jest identyfikacja „podstawowych zasad optymalności”. W artykule opublikowanym w 2018 roku na łamach czasopisma „Bioinformatics” dokonali oni przeglądu prac czołowych badaczy, którzy także dowiedli mocy predykcyjnej założenia optymalności:
William J. Sutherland14 twierdzi, że te zasady optymalności umożliwiają biologom przejście od samego wyjaśniania wzorców lub mechanizmów do możliwości formułowania przewidywań na podstawie pierwszych zasad. William Bialek15 czyni ważne spostrzeżenie, że hipotez optymalności nie należy przyjmować z powodów estetycznych, lecz jako podejście, które można bezpośrednio przetestować za pomocą eksperymentów ilościowych. Matematyczną optymalizację można więc postrzegać jako podstawowe narzędzie badawcze w bioinformatyce i obliczeniowej biologii systemów16.
Inni badacze wykazali nawet, że takie układy biologiczne, jak replikacja i translacja DNA17, rozwój zarodkowy18 i procesy sensoryczne19, działają z wydajnością bliską granic tego, co jest fizycznie możliwe. Wytwory ludzkiej inżynierii wypadają blado w porównaniu z takimi osiągnięciami przyrody.
Ogromna większość świadectw empirycznych jest zgodna z opartym na idei projektu przewidywaniem optymalności, a także bezpośrednio przeczą one głównemu przewidywaniu każdej teorii niekierowanej ewolucji. Czy te świadectwa skłonią takich krytyków, jak Kitcher i Lents, do ponownego przemyślenia swoich poglądów? Najprawdopodobniej tak się nie stanie, ponieważ podstawą dla ich przekonania o słuszności materializmu naukowego nie są dane empiryczne, lecz ich filozoficzne poglądy. Na szczęście wielu biologów nie poddaje się presji społecznej nakazującej utrzymywanie status quo i przyjmuje, że we właściwym kierunku powinny prowadzić ich dane empiryczne. Ci naukowcy przeprowadzą biologię przez kolejną wielką rewolucję naukową, która właśnie się rozpoczęła.
Brian Miller
Oryginał: Why Systems Biologists Now Assume Life Is Optimally Designed, „Evolution News & Science Today” 2021, August 27 [dostęp: 31 I 2025].
Przekład z języka angielskiego: Dariusz Sagan
Przypisy
- Por. B. Miller, Koniec drogi dla debaty o inteligentnym projekcie?, tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 24 stycznia [dostęp: 31 I 2025]; tenże, Teleologia powraca do biologii, tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 29 stycznia [dostęp: 31 I 2025].
- Argument from Poor Design, „Wikipedia”, https://en.wikipedia.org/wiki/Argument_from_poor_design [dostęp: 4 XII 2024].
- Por. D.W. Snoke, J. Cox, D. Petcher, Suboptimality and Complexity in Evolution, „Complexity” 2014, Vol. 1, No. 1, s. 322–327, https://doi.org/10.1002/cplx.21566.
- Por. S.S. Hall, Hidden Treasures in Junk DNA: What Was Once Known as Junk DNA Turns Out to Hold Hidden Treasures, Says Computational Biologist Ewan Birney, „Scientific American” 2012, October 1 [dostęp: 4 XII 2024].
- P. Kitcher, Living with Darwin: Evolution, Design, and the Future of Faith, Oxford University Press, New York 2007, s. 57 (przyp. tłum.).
- N.H. Lents, Człowiek i błędy ewolucji. Niepotrzebne kości, zepsute geny i inne niedoskonałości ludzkiego ciała, tłum. M. Zawiślak, J. Środa, Dom Wydawniczy Rebis, Poznań 2020, s. 12. W nawiasy kwadratowe ująłem podany przez autora niniejszego artykułu i przypisany Nathanowi Lentsowi fragment, który nie występuje ani w przekładzie książki Lentsa na język polski, ani w oryginale (przyp. tłum.).
- Por. J. Wells, The Receding Myth of “Junk DNA”, „Evolution News & Science Today” 2011, October 6 [dostęp: 4 XII 2024]; tenże, There You Go Again, Nathan Lents, „Evolution News & Science Today” 2018, May 11 [dostęp: 4 XII 2024]; S. Laufmann, Your “Botched Body”: Bad Design or Bad Logic?, „Evolution News & Science Today” 2018, April 18 [dostęp: 4 XII 2024]; M. Egnor, Nathan Lents Doesn’t Understand… Refrigeration, „Evolution News & Science Today” 2018, May 30 [dostęp: 4 XII 2024].
- Por. B. Miller, Denis Lamoureux on the God-of-the-Gaps Fallacy, „Evolution News & Science Today” 2018, September 28 [dostęp: 4 XII 2024].
- Por. D. Klinghoffer, Now It’s Whale Hips: Another Icon of Darwinian Evolution, Vestigial Structures, Takes a Hit, „Evolution News & Science Today” 2014, September 15 [dostęp: 4 XII 2024].
- Por. M. Günther, V. Keppler, A. Seyfarth, R. Blickhan, Human Leg Design: Optimal Axial Alignment Under Constraints, „Journal of Mathematical Biology” 2004, Vol. 48, No. 6, s. 623–646, https://doi.org/10.1007/s00285-004-0269-3; N. Sancisi, R. Caminati, V. Parenti-Castelli, Optimal Four-Bar Linkage for the Stability and the Motion of the Human Knee Prostheses, 2009 [dostęp: 4 XII 2024]; Foot Bones Explained | Foot Joints and Ankle Movements | Human Anatomy in 3D | Elearnin, „YouTube” [dostęp: 4 XII 2024]; J.-T. Seo; B.-J. Yi, Modeling and Analysis of a Biomimetic Foot Mechanism, „2009 IEEE/RSJ International Conference on Intelligent Robots and Systems” 2009, December 15, s. 1472–1477, https://doi.org/10.1109/IROS.2009.5354153.
- Por. S.C. Burgess, A.C. Etoundi, Performance Maps for a Bio-Inspired Robotic Condylar Hinge Joint, „Journal of Mechanical Design” 2014, Vol. 136, No. 11, numer artykułu: 115002, https://doi.org/10.1115/1.4028168; A.C. Etoundi, S.C. Burgess, R. Vaidyanathan, A Bio-Inspired Condylar Hinge for Robotic Limbs, „Journal of Mechanisms and Robotics” 2013, Vol. 5, No. 3, numer artykułu: 031011, https://doi.org/10.1115/1.4024471.
- Por. Evolution News, Next Phase of ENCODE Finds MORE Functional Information in Genome “Junk”, „Evolution News & Science Today” 2020, August 4 [dostęp: 4 XII 2024].
- D. Graur, Update Version of the SMBE/SESBE Lecture on ENCODE & Junk DNA (Graur, December 2013), „SlideShare” 2013, December 20 [dostęp: 4 XII 2024]. Por. też C. Luskin, Paper Shows that “Mutational Load” Arguments Don’t Refute ENCODE, „Evolution News & Science Today” 2020, May 1 [dostęp: 4 XII 2024].
- Por. W.J. Sutherland, The Best Solution, „Nature” 2005, Vol. 435, No. 7042, s. 569, https://doi.org/10.1038/435569a (przyp. tłum.).
- Por. W. Bialek, Perspectives on Theory at the Interface of Physics and Biology, „Reports on Progress in Physics” 2017, Vol. 81, No. 1, numer artykułu: 012601, https://doi.org/10.1088/1361-6633/aa995b (przyp. tłum.).
- N. Tsiantis, E. Balsa-Canto, J.R. Banga, Optimality and Identification of Dynamic Models in Systems Biology: An Inverse Optimal Control Framework, „Bioinformatics” 2018, Vol. 34, No. 14, s. 2433–2434 [2433–2440], https://doi.org/10.1093/bioinformatics/bty139.
- Por. W.D. Piñeros, T. Tlusty, Kinetic Proofreading and the Limits of Thermodynamic Uncertainty, „Physical Review E” 2019, Vol. 101, numer artykułu: 022415, https://doi.org/10.1103/PhysRevE.101.022415.
- Por. M.D. Petkova et al., Optimal Decoding of Cellular Identities in a Genetic Network, „Cell” 2019, Vol. 176, No. 4, s. 844–855.e15, https://doi.org/10.1016/j.cell.2019.01.007.
- Por. G.D. Field, A.P. Sampath, Behavioural and Physiological Limits to Vision in Mammals, „Philosophical Transactions of the Royal Society B: Biological Sciences” 2017, Vol. 372, No. 1717, numer artykułu: 20160072, https://doi.org/10.1098/rstb.2016.0072.
Literatura:
- Argument from Poor Design, „Wikipedia” [dostęp: 4 XII 2024].
- Bialek W., Perspectives on Theory at the Interface of Physics and Biology, „Reports on Progress in Physics” 2017, Vol. 81, No. 1, numer artykułu: 012601, https://doi.org/10.1088/1361-6633/aa995b.
- Burgess S.C., Etoundi A.C., Performance Maps for a Bio-Inspired Robotic Condylar Hinge Joint, „Journal of Mechanical Design” 2014, Vol. 136, No. 11, numer artykułu: 115002, https://doi.org/10.1115/1.4028168.
- Egnor M., Nathan Lents Doesn’t Understand… Refrigeration, „Evolution News & Science Today” 2018, May 30 [dostęp: 4 XII 2024].
- Etoundi A.C., Burgess S.C., Vaidyanathan R., A Bio-Inspired Condylar Hinge for Robotic Limbs, „Journal of Mechanisms and Robotics” 2013, Vol. 5, No. 3, numer artykułu: 031011, https://doi.org/10.1115/1.4024471.
- Evolution News, Next Phase of ENCODE Finds MORE Functional Information in Genome “Junk”, „Evolution News & Science Today” 2020, August 4 [dostęp: 4 XII 2024].
- Field G.D., Sampath A.P., Behavioural and Physiological Limits to Vision in Mammals, „Philosophical Transactions of the Royal Society B: Biological Sciences” 2017, Vol. 372, No. 1717, numer artykułu: 20160072, https://doi.org/10.1098/rstb.2016.0072.
- Foot Bones Explained | Foot Joints and Ankle Movements | Human Anatomy in 3D | Elearnin, „YouTube” [dostęp: 4 XII 2024].
- Graur D., Update Version of the SMBE/SESBE Lecture on ENCODE & Junk DNA (Graur, December 2013), „SlideShare” 2013, December 20 [dostęp: 4 XII 2024].
- Günther M., Keppler V., Seyfarth A., Blickhan R., Human Leg Design: Optimal Axial Alignment Under Constraints, „Journal of Mathematical Biology” 2004, Vol. 48, No. 6, s. 623–646, https://doi.org/10.1007/s00285-004-0269-3.
- Hall S.S., Hidden Treasures in Junk DNA: What Was Once Known as Junk DNA Turns Out to Hold Hidden Treasures, Says Computational Biologist Ewan Birney, „Scientific American” 2012, October 1 [dostęp: 4 XII 2024].
- Kitcher P., Living with Darwin: Evolution, Design, and the Future of Faith, Oxford University Press, New York 2007.
- Klinghoffer D., Now It’s Whale Hips: Another Icon of Darwinian Evolution, Vestigial Structures, Takes a Hit, „Evolution News & Science Today” 2014, September 15 [dostęp: 4 XII 2024].
- Laufmann S., Your “Botched Body”: Bad Design or Bad Logic?, „Evolution News & Science Today” 2018, April 18 [dostęp: 4 XII 2024].
- Lents N.H., Człowiek i błędy ewolucji. Niepotrzebne kości, zepsute geny i inne niedoskonałości ludzkiego ciała, tłum. M. Zawiślak, J. Środa, Dom Wydawniczy Rebis, Poznań 2020.
- Luskin C., Paper Shows that “Mutational Load” Arguments Don’t Refute ENCODE, „Evolution News & Science Today” 2020, May 1 [dostęp: 4 XII 2024].
- Miller B., Denis Lamoureux on the God-of-the-Gaps Fallacy, „Evolution News & Science Today” 2018, September 28 [dostęp: 4 XII 2024].
- Miller B., Koniec drogi dla debaty o inteligentnym projekcie?, tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 24 stycznia [dostęp: 31 I 2025].
- Miller B., Teleologia powraca do biologii, tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 29 stycznia [dostęp: 31 I 2025].
- Petkova M.D. et al., Optimal Decoding of Cellular Identities in a Genetic Network, „Cell” 2019, Vol. 176, No. 4, s. 844–855.e15, https://doi.org/10.1016/j.cell.2019.01.007.
- Piñeros W.D., Tlusty T., Kinetic Proofreading and the Limits of Thermodynamic Uncertainty, „Physical Review E” 2019, Vol. 101, numer artykułu: 022415, https://doi.org/10.1103/PhysRevE.101.022415.
- Sancisi N., Caminati R., Parenti-Castelli V., Optimal Four-Bar Linkage for the Stability and the Motion of the Human Knee Prostheses, 2009 [dostęp: 4 XII 2024].
- Seo J.-T.; Yi B.-J., Modeling and Analysis of a Biomimetic Foot Mechanism, „2009 IEEE/RSJ International Conference on Intelligent Robots and Systems” 2009, December 15, s. 1472–1477, https://doi.org/10.1109/IROS.2009.5354153.
- Snoke D.W., Cox J., Petcher D., Suboptimality and Complexity in Evolution, „Complexity” 2014, Vol. 1, No. 1, s. 322–327, https://doi.org/10.1002/cplx.21566.
- Sutherland W.J., The Best Solution, „Nature” 2005, Vol. 435, No. 7042, s. 569, https://doi.org/10.1038/435569a.
- Tsiantis N., Balsa-Canto E., Banga J.R., Optimality and Identification of Dynamic Models in Systems Biology: An Inverse Optimal Control Framework, „Bioinformatics” 2018, Vol. 34, No. 14, s. 2433–2440, https://doi.org/10.1093/bioinformatics/bty139.
- Wells J., The Receding Myth of “Junk DNA”, „Evolution News & Science Today” 2011, October 6 [dostęp: 4 XII 2024].
- Wells J., There You Go Again, Nathan Lents, „Evolution News & Science Today” 2018, May 11 [dostęp: 4 XII 2024].