Zarodki
Teoria ewolucji potrzebuje więc lepszego potwierdzenia empirycznego niż to zapewniane przez zapis kopalny. Coyne słusznie zauważył, że „pisząc dzieło O powstawaniu gatunków Darwin uznał embriologię za najmocniejszą podporę teorii ewolucji”1. Darwin napisał, że świadectwa empiryczne zdają się wskazywać na to, iż „zarodki bardzo różnych gatunków należących do tej samej gromady są bardzo do siebie podobne, gdy się zaś zupełnie rozwiną, stają się zupełnie różne”2, a to stanowi wzorzec, który „zdradza wspólnotę pochodzenia”3. Darwin sądził, że zarodki na wczesnym etapie rozwoju „przedstawiają mniej więcej dokładnie wygląd przodka całej grupy w jego stadium dorosłym”4.
Darwin nie był jednak embriologiem. W O powstawaniu gatunków poparł swoje twierdzenia słowami niemieckiego embriologa Karla Ernsta von Baera:
Zarodki ssaków, ptaków, jaszczurek i węży, a zapewne także żółwi, tak dalece są podobne do siebie we wczesnych stadiach rozwojowych […]. Mam dwa zarodki zachowane w alkoholu, których zapomniałem oznaczyć, a teraz zupełnie nie mogę powiedzieć, do jakiej gromady należą. Być może są to jaszczurki lub małe ptaki albo też bardzo młode ssaki, tak wielkie jest podobieństwo w sposobie powstawania głowy i tułowia u tych zwierząt5.
Coyne uważa, że są to słowa, które von Baer napisał „w liście do Darwina”6, ale Coyne zna historię równie słabo, jak paleontologię. Fragment zacytowany przez Darwina pochodzi z pracy napisanej przez von Baera po niemiecku w 1828 roku. Thomas Henry Huxley przetłumaczył tę pracę na język angielski, a przekład opublikował w 1853 roku7. Darwin z początku nawet nie zdawał sobie sprawy, że jest to wypowiedź von Baera: w pierwszych dwóch wydaniach O powstawaniu gatunków błędnie przypisywał ją Louisowi Agassizowi8.
Jak na ironię, von Baer był zdecydowanym krytykiem teorii Darwina i odrzucał pogląd, że kręgowce mają wspólnego przodka. Jak twierdzi historyk nauki Timothy Lenoir, von Baer obawiał się, iż Darwin i jego następcy „uznali, że darwinowska hipoteza ewolucji jest prawdziwa, zanim jeszcze podjęli się obserwacji zarodków”9. Mit, zgodnie z którym praca von Baera wspierała teorię Darwina, rozpowszechnił głównie inny niemiecki biolog – Ernst Haeckel10. Haeckel utrzymywał nie tylko, że wszystkie zarodki kręgowców pochodzą od wspólnego przodka, ale też że proces ich rozwoju („ontogeneza”) odtwarza („rekapituluje”) ich historię ewolucyjną („filogenezę”). Nazwał to prawem biogenetycznym: ontogeneza jest rekapitulacją filogenezy.
W książce Ewolucja jest faktem Coyne napisał, że „podobną «rekapitulację» sekwencji ewolucyjnej można również dostrzec w rozwoju”11 różnych narządów. „Każde zwierzę kręgowe przechodzi przez kolejne etapy rozwoju i tak się składa, że sekwencja tych etapów naśladuje ewolucyjną sekwencję jego przodków”12. Prawdopodobnym tego powodem jest to, że „kiedy jeden gatunek przeistacza się w inny, jego potomkowie dziedziczą program rozwojowy swoich przodków”13. Tak więc z perspektywy potomka „korzystniej jest posługiwać się gotowym i sprawdzonym planem rozwoju, modyfikując tylko nieznacznie poszczególne struktury. Lepiej, żeby cechy, które wyewoluowały później w filogenezie, rozwijały się też później w ontogenezie. Ta zasada «dokładania nowych cegiełek do starej budowli» tłumaczy również, dlaczego sekwencja zmian rozwojowych naśladuje ewolucyjną sekwencję gatunków. Kiedy nowy gatunek powstaje ze starego, zachowuje program rozwojowy odziedziczony po swym przodku, uzupełniając go jedynie na końcu o nowe dodatki”14. Dlatego „wszystkie kręgowce na początku swego rozwoju wyglądają jak zarodki ryb”15.
Niemniej według Coyne’a Prawo Biogenetyczne Haeckela „nie jest do końca prawdziwe”16, ponieważ „kolejne stadia w rozwoju zarodkowym gatunku nie przypominają wcale dorosłych form jego przodków”, jak sądził Haeckel, „ale zarodki tych przodków”17. To przeformułowanie Prawa Biogenetycznego nie rozwiązuje jednak problemu. Po pierwsze, skamieniałe zarodki są zjawiskiem skrajnie rzadkim18, a więc podstawą dla tego przeformułowanego prawa są wyłącznie zarodki współczesnych organizmów, co do których zakłada się, że przypominają formy ancestralne. Otrzymujemy więc argument oparty na błędnym kole: skoro zgodnie z teorią Darwina ryby są naszymi przodkami, a ludzkie zarodki (rzekomo) przypominają zarodki ryb, zatem ludzkie zarodki wyglądają jak zarodki naszych przodków. Teoria ma pierwszeństwo, a obserwacje grają drugorzędną rolę – dokładnie taki zarzut postawił von Baer.
Po drugie, idea, że późniejsze stadia ewolucyjne można powiązać z procesem rozwoju, jest nierealistyczna pod względem biologicznym. Człowiek nie jest jedynie zarodkiem ryby z dodatkiem jakichś innych cech. Jak w 1922 roku wykazał brytyjski embriolog Walter Garstang, „dom nie jest po prostu rozbudowaną chatą. Stanowi on wyższy poziom w ewolucji miejsca zamieszkania, ale cały budynek został przekształcony – fundamenty, drewniane belki i dach – nawet jeżeli wykorzystano te same cegły”19.
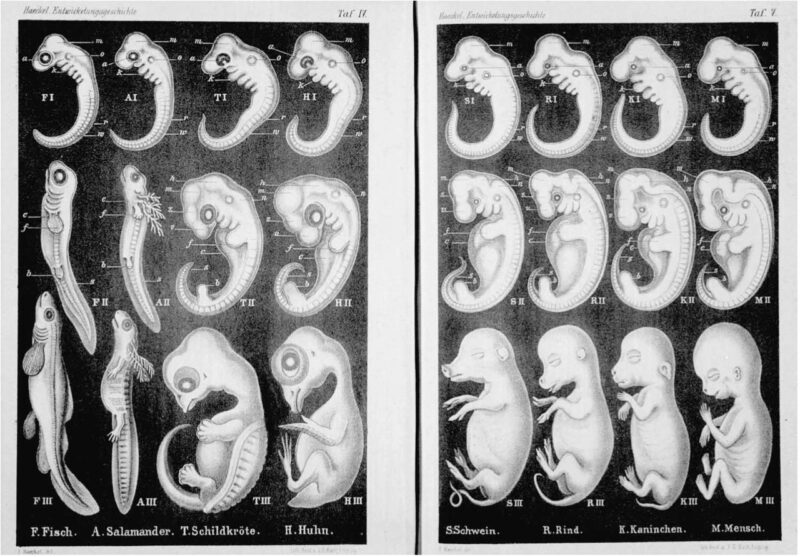
Po trzecie i najważniejsze, zarodki kręgowców nie są najbardziej podobne do siebie na najwcześniejszych etapach. W latach 1860-tych Haeckel sporządził rysunki w celu pokazania, że zarodki kręgowców wyglądają niemal identycznie w pierwszym stadium rozwoju. Rysunki te były jednak sfałszowane. Haeckel nie tylko zniekształcił zarodki tak, aby bardziej przypominały siebie nawzajem niż jest w rzeczywistości, ale też pominął wcześniejsze stadia, w których zarodki są uderzająco odmienne od siebie. Ludzki zarodek w swoich najwcześniejszych stadiach w ogóle nie przypomina zarodka ryby.
Dopiero w połowie procesu rozwoju ludzkie zarodki osiągają stadium, które Darwin i Haeckel uznali za pierwsze. Ten wzorzec różnicy-podobieństwa-różnicy nazywany jest przez specjalistów w dziedzinie biologii rozwoju „klepsydrą rozwojową”. Zarodki kręgowców nie przypominają siebie nawzajem w swoich najwcześniejszych stadiach, ale są do siebie podobne w środkowym stadium rozwoju, a potem znowu pojawiają się między nimi różnice. Gdyby ontogeneza była rekapitulacją filogenezy, taki wzorzec byłby bardziej spójny z poglądem o oddzielnym powstaniu niż z teorią wspólnoty pochodzenia. Współcześni darwiniści próbują uchronić swoją teorię dzięki przyjęciu założenia, że wspólnota pochodzenia kręgowców zostaje przesłonięta z powodu łatwego ewoluowania takich samych wczesnych stadiów procesu rozwoju. Założenie to nie ma jednak niezależnego uzasadnienia – wynika ono wyłącznie z teorii ewolucji20.
Chociaż już w czasach Haeckela wykazano, że jego rysunki są sfałszowane, to przez cały wiek dwudziesty powielano je w podręcznikach biologii po to, aby przekonać studentów, że ludzie mają wspólnego przodka z rybami. Później, w 1997 roku, w jednym z czasopism naukowych opublikowano artykuł, w którym porównano zdjęcia zarodków kręgowców z rysunkami Haeckela21. Główny autor tego artykułu stwierdził, że te rysunki stanowią „jedno z najsłynniejszych fałszerstw w biologii”22. W 2000 roku harwardzki biolog ewolucyjny Stephen Jay Gould nazwał rysunki Haeckela „oszustwem” i stwierdził, że biologowie powinni być „zarówno zdumieni, jak i zawstydzeni tym, że przez sto lat bezmyślnie powielano i utrwalano te rysunki w wielu, a może nawet w większości współczesnych podręczników”23.
Coyne nie czuje jednak wstydu i wręcz broni rysunków Haeckela: „Prawo biogenetyczne popadło z czasem w niełaskę, nie tylko dlatego, że nie jest do końca prawdziwe, ale również z powodu oskarżeń, też nie całkiem sprawiedliwych, że Haeckel zafałszował na swych rysunkach rzeczywistość, znacznie przesadzając z podobieństwem wszystkich wczesnych form rozwojowych kręgowców. Nie powinniśmy jednak wylewać dziecka razem z kąpielą”24. „Dzieckiem” jest teoria Darwina, której Coyne uparcie broni, nie zważając na dane empiryczne.
Narządy szczątkowe i nieudany projekt
W O powstawaniu gatunków Darwin argumentował, że powszechne występowanie narządów szczątkowych – które mogły pełnić kiedyś funkcję, ale teraz są bezużyteczne – stanowi świadectwo przeczące idei stworzenia: „Jeślibyśmy przyjęli, iż każdy organizm został oddzielnie stworzony wraz ze wszystkimi swymi poszczególnymi częściami, jakże niezrozumiały stałby się dla nas fakt, że narządy, noszące na sobie wyraźnie piętno nieużyteczności […] występują tak często”25. Darwin przekonywał jednocześnie, że takie narządy z łatwością wyjaśnia jego teoria: „Przyjmując teorię wspólnoty pochodzenia i stopniowych przekształceń możemy stwierdzić, że istnienie narządów w stanie szczątkowym, niedoskonałym i nieużytecznym lub też zupełny ich zanik wcale nie są tak trudne do wyjaśnienia, jak bez wątpienia były trudne w oparciu o starą teorię stworzenia i że nawet zjawisko to można by było przewidzieć na podstawie wyłożonych tu poglądów”26.
W książce O pochodzeniu człowieka Darwin uznał, że przykładem narządu szczątkowego jest ludzki wyrostek robaczkowy27. Był on jednak w błędzie: wiemy obecnie, że wyrostek robaczkowy stanowi ważne źródło krwinek produkujących przeciwciała, a w związku z tym jest integralną częścią ludzkiego układu odpornościowego. Może też pełnić funkcję zbiornika na korzystne dla nas bakterie potrzebne do normalnego przebiegu procesu trawienia. Okazuje się więc, że wyrostek robaczkowy wcale nie jest bezużyteczny28.
W 1981 roku kanadyjski biolog Steven Scadding zapewniał, że wprawdzie nie ma on żadnych obiekcji względem darwinizmu, ale jego zdaniem „narządy szczątkowe nie stanowią świadectwa na rzecz teorii ewolucji”29. Główny tego powód polega na tym, że „jednoznaczne rozstrzygnięcie, czy narząd nie pełni żadnej funkcji, jest trudne, a być może nawet niemożliwe”30. Jako przykład Scadding podał wyrostek robaczkowy, który uznawano niegdyś za narząd szczątkowy, ale obecnie wiemy, że jest on funkcjonalny. Inny kanadyjski biolog Bruce Naylor odpowiedział, że narząd pełniący jakąś funkcję wciąż można uznawać za szczątkowy. Argumentował on też, że „doskonale zaprojektowane organizmy wymagałyby istnienia stwórcy”, ale „organizmy często nie są doskonale zaprojektowane”31 i fakt ten lepiej wyjaśnia teoria ewolucji. Scadding odparł natomiast, że „cała siła argumentu Darwina i innych biologów, dotyczącego narządów szczątkowych, tkwi w tym, że są one bezużyteczne i bezcelowe”32. W przeciwnym razie argument dotyczący narządów szczątkowych staje się jedynie argumentem z homologii, a „Darwin oddzielał te argumenty od siebie, ponieważ jego zdaniem są one wzajemnie niezależne”33. Scadding zarzucił ponadto Naylorowi, że argument mówiący o „niedoskonałym zaprojektowaniu” opiera się na „teologicznym założeniu na temat natury Boga, mianowicie że nie stworzyłby on bezużytecznych struktur. Niezależnie od tego, czy to teologiczne twierdzenie jest zasadne, bez wątpienia nie można uznać, że ma ono charakter naukowy, a tym samym nie powinno być ono przywoływane w naukowej debacie na temat ewolucji”34.
W książce Ewolucja jest faktem Coyne (podobnie jak Darwin) twierdzi, że ludzki wyrostek robaczkowy jest przykładem narządu szczątkowego. Jednakże w odróżnieniu od Darwina, przyznaje on, że „ten nasz wyrostek pełni jednak pewną, choć bardzo niepozorną, funkcję. W jego wnętrzu znajdują się wiązki tkanek, które mogą stanowić część układu odpornościowego. Są też sugestie, że daje on schronienie ważnym bakteriom jelitowym, kiedy w wyniku infekcji giną one w pozostałej części układu pokarmowego. Niemniej te drobne korzyści bledną w obliczu poważnych problemów, które nasz wyrostek wywołuje”35. Coyne stwierdza dalej: „Jeśli położymy na szali z jednej strony te niewielkie korzyści, jakie mamy z wyrostków robaczkowych, i wielkie szkody, jakie mogą one powodować, jasne jest, że lepiej, by ich w ogóle nie było. Ale są, choć w formie szczątkowej, i z pewnością nie pełnią już swych pierwotnych funkcji”36.
Jak jednak niemal trzydzieści lat temu wskazywał Scadding, siła argumentu Darwina polega na fakcie niefunkcjonalności narządu. Co więcej, gdyby szczątkowość przedefiniowano zgodnie z propozycją Coyne’a, obejmowałaby ona wiele cech, których do tej pory nie uznawano za szczątkowe. Na przykład, jeżeli ludzka ręka jest przekształconą nogą czworonożnego ssaka (jak twierdzą darwiniści), to przy takiej definicji jest ona narządem szczątkowym. I jeśli (jak argumentuje Coyne) skrzydła ptaków zdolnych do latania wyewoluowały z upierzonych kończyn przednich dinozaurów, które używały ich do innych celów, to te skrzydła również mają charakter szczątkowy37. Większość ludzi inaczej rozumie jednak pojęcie „szczątkowości”.
Coyne ignoruje też pozostałe zarzuty Scaddinga, argumentując, że niezależnie od tego, czy ludzki wyrostek robaczkowy jest bezużyteczny czy jednak pełni jakąś funkcję, stanowi on przykład niedoskonałego lub nieudanego projektu. Coyne pisze tak: „Używając określenia «nieudany projekt», mam na myśli to, że gdyby organizmy były zaprojektowane od zera przez inteligentnego projektanta – z tych samych biologicznych materiałów (nerwy, mięśnie, kości i tak dalej) – nie wykazywałyby tych wszystkich niedoskonałości. Projekt doskonały byłby czytelnym znakiem firmowym inteligentnego i wytrawnego projektanta. To właśnie niedoskonały projekt przemawia za ewolucją – co więcej, tego właśnie od ewolucji oczekujemy”38.
Według Coyne’a jeszcze lepszy przykład nieudanego projektu stanowią powszechnie występujące „martwe geny”. Zgodnie ze współczesną odmianą darwinizmu, której broni Coyne, w DNA zawarty jest program genetyczny kodujący białka regulujące proces rozwoju zarodka. Mutacje niekiedy zmieniają program genetyczny, prowadząc do produkcji nowych białek (lub zmiany ich położenia). Następnie dobór naturalny oddziałuje na rezultaty tych mutacji i tak zachodzi ewolucja. W latach 1970-tych biologowie molekularni odkryli jednak, że większość naszego DNA nie koduje białek. W 1972 roku Susumu Ohno określił takie DNA mianem „śmieciowego”39. W 1976 roku Richard Dawkins napisał: „znaczna część DNA nie jest nigdy tłumaczona na białko. Z punktu widzenia pojedynczego organizmu wydaje się to paradoksalne. Jeśli «przeznaczeniem» DNA jest nadzór nad budowaniem ciał, zaskakujące jest odkrycie, że znaczna jego część nie uczestniczy w tym dziele”40. Z perspektywy ewolucjonizmu darwinowskiego nie ma w tym jednak nic paradoksalnego: „Rzeczywistym «celem» DNA jest przetrwanie – tyle i nic ponadto. Najprościej można uzasadnić obecność nadmiaru DNA, przyjmując, że jest on pasożytem lub jedynie pasażerem na gapę – nieszkodliwym, ale bezużytecznym, który zabrał się maszyną przetrwania stworzoną przez inny DNA”41.
Tak samo jak Dawkins, Coyne uważa, że duża część naszego DNA jest pasożytem. W książce Ewolucja jest faktem Coyne napisał: „gdy jakaś cecha przestaje być użyteczna lub zostaje zredukowana, stojące za nią geny nie znikają od razu z genomu: ewolucja je wyłącza, ale nie wymazuje ich zupełnie z DNA. To pozwala nam wysunąć pewne przypuszczenie – takie mianowicie, że w genomach wielu gatunków istnieją wyłączone albo «martwe» geny, które kiedyś były aktywne, ale dziś są już zdegradowane i nie podlegają ekspresji. Innymi słowy, geny szczątkowe. Gdyby wszystkie gatunki były stworzone z niczego, należałoby przypuszczać, że takie geny (i narządy) nie powinny istnieć”42. Dalej Coyne twierdzi, że „trzydzieści lat temu nie moglibyśmy sprawdzić tych przypuszczeń, bo nie znaliśmy sposobu odczytywania kodu genetycznego. Dzisiaj jednak możemy bardzo łatwo sekwencjonować pełne genomy różnych gatunków i dla wielu, w tym dla człowieka, taki odczyt został dokonany. A to daje nam do ręki wyjątkowo precyzyjne narzędzie do badań nad ewolucją, jeśli będziemy pamiętać, że normalną funkcją genu jest kodowanie białek, których sekwencja aminokwasów określona jest przez sekwencję zasad nukleinowych budujących DNA. Kiedy mamy więc sekwencję DNA danego genu, możemy zwykle ustalić, czy podlega on normalnej ekspresji – czy wytwarza funkcjonalne białko – czy też został uciszony i niczego już nie wytwarza. Możemy na przykład stwierdzić, czy mutacje tak bardzo uszkodziły gen, że nie jest już w stanie wytwarzać funkcjonalnych białek, czy może wyłączone zostały obszary «kontrolne» genu, odpowiedzialne za jego aktywację. Gen, który przestał już funkcjonować, nazywany jest pseudogenem. Nasza ewolucyjna predykcja, że powinniśmy znajdować w genomie pseudogeny, została potwierdzona – i to wielokrotnie. Praktycznie każdy gatunek zawiera martwe geny, z których wiele ma aktywne odpowiedniki u gatunków pokrewnych. A to oznacza, że geny te były również czynne u wspólnego przodka i zostały wyłączone w niektórych – ale nie wszystkich – liniach potomnych. Na przykład spośród około 30 000 genów ludzkiego genomu ponad dwa tysiące to pseudogeny. Nasz genom – i genomy innych gatunków – to cmentarzyska martwych genów”43.
Coyne jest jednak w grubym błędzie.
Dane empiryczne uzyskiwane w ramach projektów sekwencjonowania genomów wskazują, że praktycznie całe DNA danego organizmu ulega transkrypcji na RNA i że chociaż większość RNA nie jest tłumaczona na białka, to pełni on istotne funkcje regulacyjne. Każdego miesiąca w czasopismach naukowych publikowane są artykuły opisujące coraz więcej takich funkcji. Nie są to wiadomości z ostatniej chwili: od 2003 roku (kiedy naukowcy zsekwencjonowali genom człowieka) gromadzono dane wskazujące, że „pseudogeny” i inne tak zwane sekwencje „śmieciowego DNA” wcale nie są bezużyteczne44. W książce Ewolucja jest faktem Coyne ignoruje ten ogromny zespół danych, które bezapelacyjnie obalają darwinowskie przewidywanie, że nasz genom powinien zawierać duże ilości „martwego” DNA. Nie ma się co dziwić, że Coyne ciągle stosuje argumenty teologiczne, które według Scaddinga „nie powinny być przywoływane w naukowej debacie na temat ewolucji”45.
Oryginał: Why Darwinism Is False, „Discovery Institute” 2009, May 18 [dostęp: 7 VI 2024].
Przekład z języka angielskiego: Dariusz Sagan
{kind=link}
Przypisy
- J. Coyne, Ewolucja jest faktem, s. 105.
- K. Darwin, O powstawaniu gatunków, s. 508.
- Tamże, s. 518.
- Tamże, s. 517–518.
- Cyt. za: tamże, s. 508–509.
- J. Coyne, Ewolucja jest faktem, s. 98.
- Por. K.E. von Baer, On the Development of Animals, with Observations and Reflections: The Fifth Scholium, trans. T.H. Huxley, w: Scientific Memoirs: Selected from the Transactions of Foreign Academies of Science and from Foreign Journals: Natural History, eds. A. Henfrey, T.H. Huxley, Taylor and Francis, London 1853 (przedruk: New York 1966), s. 186–237. Fragment zacytowany przez Darwina znajduje się na s. 210.
- Por. J.M. Oppenheimer, An Embryological Enigma in the Origin of Species, w: J.M. Oppenheimer, Essays in the History of Embryology and Biology, MIT Press, Cambridge 1967, s. 221–255.
- T. Lenoir, The Strategy of Life: Teleology and Mechanics in Nineteenth Century German Biology, University of Chicago Press, Chicago 1982, s. 258.
- Por. F.B. Churchill, The Rise of Classical Descriptive Embryology, w: Developmental Biology: A Comprehensive Synthesis, Vol. 7: A Conceptual History of Modern Embryology, ed. S.F. Gilbert, Plenum Press, Baltimore 1991, s. 19–20 [1–29].
- J. Coyne, Ewolucja jest faktem, s. 102.
- Tamże, s. 103.
- Tamże.
- Tamże [wyróżnienia w oryginale].
- Tamże, s. 104.
- Tamże.
- Tamże [wyróżnienie w oryginale].
- Por. S. Conway Morris, Fossil Embryos, w: Gastrulation: From Cells to Embryos, ed. C.D. Stern, Harbour Laboratory Press, Cold Spring Harbor 2004, s. 703–711.
- W. Garstang, The Theory of Recapitulation: A Critical Restatement of the Biogenetic Law, „Journal of the Linnean Society (Zoology)” 1922, Vol. 35, s. 81–101.
- Por. rozdział piąty mojej książki Ikony ewolucji, a także rozdział trzeci mojej książki The Politically Incorrect Guide to Darwinism and Intelligent Design [Politycznie niepoprawny przewodnik po darwinizmie i teorii inteligentnego projektu] oraz podane w nich odnośniki bibliograficzne.
- Por. M.K. Richardson et al., There Is No Highly Conserved Embryonic Stage in the Vertebrates: Implications for Current Theories of Evolution and Development, „Anatomy & Embryology” 1997, Vol. 196, No. 2, s. 91–106, https://doi.org/10.1007/s004290050082.
- Cytat z M.K. Richardsona por. w: E. Pennisi, Haeckel’s Embryos: Fraud Rediscovered, „Science” 1997, Vol. 277, No. 5331, s. 1435, https://doi.org/10.1126/science.277.5331.1435a.
- S.J. Gould, Abscheulich! Atrocious!, „Natural History” 2000 (March), s. 42–49. Por. też Hoax of Dodos 2007 [dostęp: 1 VI 2024].
- J. Coyne, Ewolucja jest faktem, s. 104.
- K. Darwin, O powstawaniu gatunków, s. 550.
- Tamże, s. 526.
- Por. K. Darwin, O pochodzeniu człowieka. Dzieła wybrane, t. IV, tłum. S. Panek, „Biblioteka Klasyków Biologii”, Państwowe Wydawnictwo Rolnicze i Leśne, Warszawa 1959, s. 20.
- Por. K. Fujihashi et al., Human Appendix B Cells Naturally Express Receptors for and Respond to Interleukin 6 with Selective IgA1 and IgA2 Synthesis, „Journal of Clinical Investigations” 1991, Vol. 88, s. 248–252, https://doi.org/10.1172/JCI115284; J.A. Laissue et al., The Intestinal Immune System and Its Relation to Disease, „Digestive Diseases (Basel)” 1993, Vol. 11, No. 4–5, s. 298–312, https://doi.org/10.1159/000171421; L.G. Martin, What is the Function of the Human Appendix? Did It Once Have a Purpose That Has Since Been Lost?, „Scientific American” 1999, October 21 [dostęp: 1 VI 2024]; R.R. Bollinger et al., Biofilms in the Large Bowel Suggest an Apparent Function of the Human Vermiform Appendix, „Journal of Theoretical Biology” 2007, Vol. 249, No. 4, s. 826–831, https://doi.org/10.1016/j.jtbi.2007.08.032; Duke University Medical Center, Appendix Isn’t Useless At All: It’s A Safe House For Good Bacteria, „Science Daily” 2007, October 8 [dostęp: 1 VI 2024].
- S.R. Scadding, Do “Vestigial Organs” Provide Evidence for Evolution?, „Evolutionary Theory” 1981, Vol. 5, s. 173–176.
- Tamże.
- B.G. Naylor, Vestigial Organs Are Evidence of Evolution, „Evolutionary Theory” 1982, Vol. 6, s. 91–96.
- S.R. Scadding, Vestigial Organs Do Not Provide Scientific Evidence for Evolution, „Evolutionary Theory” 1982, Vol. 6, s. 171–173.
- Tamże.
- Tamże.
- J. Coyne, Ewolucja jest faktem, s. 85–86.
- Tamże, s. 86.
- Por. tamże, s. 66.
- Tamże, s. 107 [wyróżnienia w oryginale].
- Por. S. Ohno, So Much «Junk» DNA in Our Genome, „Brookhaven Symposia in Biology” 1972, Vol. 23, s. 366–370.
- R. Dawkins, Samolubny gen, tłum. M. Skoneczny, Prószyński i S-ka, Warszawa 1996, s. 72.
- Tamże, s. 73.
- J. Coyne, Ewolucja jest faktem, s. 91.
- Tamże, s. 91–92 [wyróżnienie w oryginale].
- Oto kilka z wielu artykułów naukowych opublikowanych po 2003 roku, w których opisuje się funkcje tak zwanego „śmieciowego” DNA: E.S. Balakirev, F.J. Ayala, Pseudogenes: Are They «Junk» or Functional DNA?, „Annual Review of Genetics” 2003, Vol. 37, s. 123–151, https://doi.org/10.1146/annurev.genet.37.040103.103949; A. Hüttenhofer, P. Schattner, N. Polacek, Non-Coding RNAs: Hope or Hype?, „Trends In Genetics” 2005, Vol. 21, No. 5, s. 289–297, https://doi.org/10.1016/j.tig.2005.03.007; J.S. Mattick, I.V. Makunin, Non-Coding RNA, „Human Molecular Genetics” 2006, Vol. 15, No. 1, s. R17–R29, https://doi.org/10.1093/hmg/ddl046; R.K. Slotkin, R. Martienssen, Transposable Elements and the Epigenetic Regulation of the Genome, „Nature Reviews Genetics” 2007, Vol. 8, s. 272–285, https://doi.org/10.1038/nrg2072; P. Carninci, J. Yasuda, Y. Hayashizaki, Multifaceted Mammalian Transcriptome, „Current Opinion in Cell Biology” 2008, Vol. 20, No. 3, s. 274–280, https://doi.org/10.1016/j.ceb.2008.03.008; C.D. Malone, G.J. Hannon, Small RNAs as Guardians of the Genome, „Cell” 2009, Vol. 136, No. 4, s. 656–668, https://doi.org/10.1016/j.cell.2009.01.045; C.P. Ponting, P.L. Oliver, W. Reik, Evolution and Functions of Long Noncoding RNAs, „Cell” 2009, Vol. 136, No. 4, s. 629–641, 10.1016/j.cell.2009.02.006.
- S.R. Scadding, Vestigial Organs Do Not Provide Scientific Evidence for Evolution.
Otóż to! W książce „Co kryje się w naszych genach?” Prof. Ewa Bartnik pisze:
(pogrubienia moje)