
Niniejszy tekst stanowi piątą część1 w serii artykułów Davida Ussery’ego zatytułowanej Czy istnieje granica ewolucji? Autor tę serię artykułów adresuje zarówno do tych, którzy jeszcze nie przeczytali książki Behego, jak i do tych, którzy już ją znają. Ussery szczegółowo analizuje argumenty przedstawione w książce Michaela J. Behego Granica ewolucji. Uważnie przygląda się pierwszym dziewięciu rozdziałom publikacji Behego oraz wskazuje mocne i słabe strony zawartych w nich argumentów. Krótko przedstawia również swoje osobiste i filozoficzne poglądy.
Rozdział 8 – Zarzuty pod adresem koncepcji granicy
Zgadzam się z Michaelem J. Behem, gdy w tym rozdziale książki Granica ewolucji. W poszukiwaniu ograniczeń darwinizmu2 pisze: „Pamiętajmy, że to nie czas jest głównym czynnikiem procesu ewolucji – jest nim liczebność populacji”3. W wyjaśnieniu tego powinna pomóc pewna analogia. W mojej dziedzinie badania prowadzone są w dużej mierze przy wykorzystaniu komputerów. Załóżmy na przykład, że mam przeprowadzić analizę porównawczą tysiąca sekwencji genomów bakteryjnych. Jeśli analiza jednego genomu zajmuje kilka dni, to analiza tysiąca genomów zajmie dosłownie LATA. Jak można ten problem obejść? Trzeba równocześnie użyć wielu procesorów. Jeśli dysponujemy tysiącem procesorów, to w zasadzie – przy założeniu, że te procesory są dostępne i wszystko idzie zgodnie z planem – jesteśmy w stanie przeprowadzić całą analizę w ciągu kilku dni. Dzięki równoczesnemu wykorzystaniu wielu procesorów można więc ogromnie przyspieszyć cały proces. To samo dotyczy procesu ewolucji. Chociaż częstość mutacji może być mała, to przy dostatecznej liczbie genomów szanse na uzyskanie „właściwej kombinacji” znacznie rosną, zwłaszcza jeśli idzie to w parze ze sporadyczną rekombinacją genomów.
W tym rozdziale Behe argumentuje w zasadzie, że nawet przy uwzględnieniu kilkuset milionów lat ewolucji nie ma dostatecznej ilości czasu na zajście „właściwych mutacji”, by w wyniku „losowego procesu” mogła wyewoluować obserwowana przez nas złożoność roślin i zwierząt. Na stronie 185 Behe stawia następujące pytanie: „Skoro tak niewiele to daje, to dlaczego mechanizm losowych mutacji i doboru naturalnego jest tak wysoko ceniony przez biologów?”4. Następnie porównuje on ideę losowych mutacji do idei „eteru”, czyli tajemniczej substancji, której istnienie postulowano ponad sto lat temu. Koncepcję tę całkowicie podważył później Albert Einstein. To porównanie jasno wskazuje na to, że w przekonaniu Behego „losowe mutacje” są mitem przyjmowanym przez większość biologów na wiarę i mającym niewielkie potwierdzenie w danych empirycznych.
Nie zgadzam się. Wierzę, że historia życia jest przepełniona celowością i że odpowiada za to Bóg. Z mojego punktu widzenia należy pytać tutaj nie o to, czy istnieje celowość, lecz o to, czy przedstawione w książce Behego argumenty naukowe mają sens i są zasadne, biorąc pod uwagę aktualną wiedzę w dziedzinach biochemii i biologii molekularnej. To właśnie do tych argumentów się tutaj odniosę. Aby naprawdę zrozumieć potencjał mutacji do tworzenia nowych interakcji między białkami, trzeba dostrzec szerszy obraz niż ten rysowany przez Behego. Bakterie pojawiły się w pierwszych ekosystemach, czyli ponad 4 miliardy lat temu, i nadal stanowią najpowszechniejszą formę życia na naszej planecie. Dysponuję tabelą, którą uwielbiam pokazywać studentom w trakcie zajęć. Pochodzi ona z artykułu przeglądowego opublikowanego rok temu5 [z perspektywy 2010 roku]. Na Ziemi istnieje 1031 bakteriofagów (wirusów atakujących bakterie) i gdybyśmy całkowicie rozciągnęli ich genomy, to miałyby one długość około tysiąca razy większą od długości Drogi Mlecznej! Gdybyśmy z naszej planety rozciągnęli cały bakteryjny DNA, miałby on długość bliską MILION razy większą niż długość Drogi Mlecznej! Jest to więc ogromna ilość DNA. Ponieważ bakterie żyją bardzo krótko (niecały dzień), więc codziennie replikacji ulega jeszcze większa ilość DNA. Każda replikacja daje okazję do nastąpienia zmian genetycznych w równoległych liniach, a zmiany genetyczne zapewniają okazję do częstego mieszania i dopasowywania się w historii życia. Nawet badając bardzo niewielki ułamek tej całości – „zaledwie” tysiąc bakteryjnych genomów – niezwykła różnorodność absolutnie wprawia mnie w osłupienie. Ani jedno białko nie jest utrwalone nawet w tej maleńkiej próbce bakterii, które zbadaliśmy do tej pory, a u wielu „gatunków” bakterii mniej niż połowa białek w jednym genomie występuje również w innym genomie – tego samego gatunku! W jakim stopniu Behe uwzględnia ten ogromny potencjał do tworzenia nowych kombinacji białek?
Behe dochodzi do zdumiewającego wniosku. Stwierdza, że „powstanie nawet jednego użytecznego wewnątrzkomórkowego miejsca wiązania między białkami może znajdować się poza zasięgiem losowych mutacji”6. Zacznijmy od sprawdzenia literatury. W bazie danych PubMed trzeba przeszukać ponad 20 milionów artykułów. Jeśli wpiszemy słowa „evolution, protein binding sites”, to znajdziemy artykuł zatytułowany Structural Features and Evolution of Protein-Protein Interactions7 [Strukturalne cechy i ewolucja interakcji między białkami] oraz 5400 innych tekstów na ten temat. W abstrakcie tej wymienionej z tytułu publikacji znajduje się następujące zdanie:
W niniejszym artykule przeanalizowano 750 krótkotrwałych interakcji między białkami oraz 2000 interakcji między domenami tego samego łańcucha białkowego (interakcje obligatoryjne) w celu lepszego zrozumienia procesu rozpoznawania na poziomie molekularnym oraz zidentyfikowania cech umożliwiających przewidywanie miejsc wiązania między białkami8.
A to tylko jeden artykuł. Czy zgodziłbyś się, że twierdzenie Behego, iż „powstanie nawet jednego użytecznego wewnątrzkomórkowego miejsca wiązania między białkami może znajdować się poza zasięgiem losowych mutacji”, najprawdopodobniej nie ma zbyt wielkiego sensu? Wydaje się, że Behe podsumowanie stanu badań biologicznych zaprezentował na forum publicznym przedwcześnie. Bez wątpienia obraz zarysowany przez Behego jest bardzo niekompletny. Zachęcam zwłaszcza do przeczytania znakomitego artykułu Kelsey Luomy na ten temat9. Luoma to studentka studiów licencjackich, która uczyniła to, co robią wszyscy dobrzy studenci na kierunkach przyrodniczych: dokonała przeglądu literatury – i okazało się, że w literaturze wyraźnie wykazano zachodzenie ewolucji nowych interakcji między białkami.
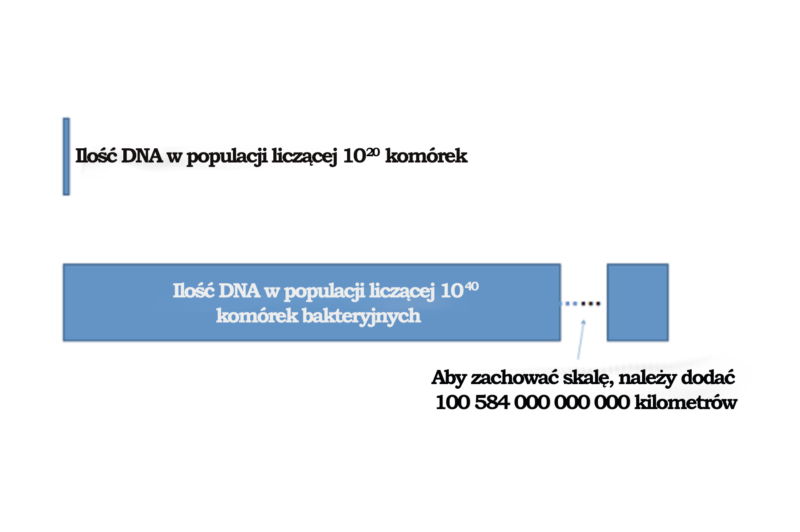
Behe ewidentnie jest więc w błędzie, gdy na stronie 175 pisze, że ponieważ „nie zaobserwowaliśmy żadnych nowych interakcji między białkami w populacji liczącej 1020 komórek, możemy z całkiem dużą pewnością wnioskować, że (co najmniej) żadne nowe układy komórkowe wymagające dwóch nowych interakcji między białkami nie pojawią się w populacji liczącej 1040 komórek – czyli w całej historii życia na Ziemi”10. W zależności od znajomości matematyki można pomyśleć, że różnica między liczbami 1020 a 1040 nie jest aż tak duża. Na wszelki wypadek przedstawmy więc tę różnicę za pomocą ilustracji. DNA z populacji liczącej 1020 komórek bakteryjnych rozciągałoby się na około 18 lat świetlnych – to duża ilość DNA! Jednak długość DNA wszystkich bakterii aktualnie żyjących na naszej planecie (w przybliżeniu 1031) wynosiłaby około 100 000 000 000 LAT ŚWIETLNYCH! A to przecież tylko ilość aktualnie istniejącego DNA bakteryjnego. Bakterie rozmnażają się bardzo szybko – raz na każde pięć minut. W porównaniu z DNA zawartym w populacji liczącej 1020 komórek (18 lat świetlnych) ilość DNA w populacji liczącej 1040 komórek wynosiłaby więc niemal 1 800 000 000 000 000 000 000 lat świetlnych. To ogromna ilość DNA. (Pamiętajmy, że jedna sekunda roku świetlnego odpowiada prawie 290 000 kilometrów. To duża ilość DNA). Bezpieczniej jest więc nie mówić na forum publicznym, że „możemy z całkiem dużą pewnością wnioskować, że (co najmniej) żadne nowe układy komórkowe wymagające dwóch nowych interakcji między białkami nie pojawią się w populacji liczącej 1040 komórek – czyli w całej historii życia na Ziemi”. Wytworzenie takiej ilości DNA zapewnia mnóstwo okazji do utworzenia nowych interakcji między białkami wskutek mutacji.
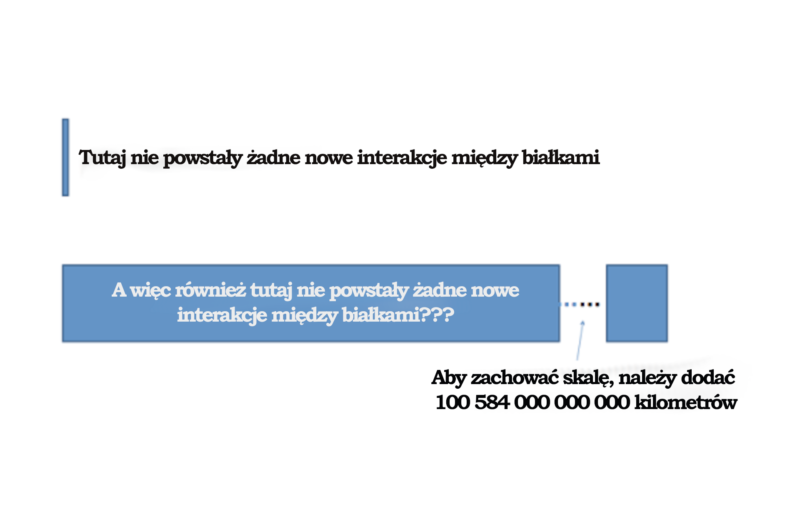
Przyjrzyjmy się bliżej, czego dotyczył eksperyment z wykorzystaniem populacji liczącej 1020 komórek, do którego odnosi się Behe w cytowanej wyżej wypowiedzi ze strony 175 jego książki, gdzie wyraźnie twierdzi on, że nie zaobserwowano żadnych nowych interakcji między białkami. W ramach tego eksperymentu badacze w istocie nie analizowali proteomu w poszukiwaniu nowych interakcji między białkami – interesował ich tylko jeden konkretny typ mutacji. Behe nie tylko dokonał więc ekstrapolacji z próbki o wielkości główki szpilki (populacji liczącej 1020 komórek) na próbkę (porównywalnie) większą od Wszechświata, lecz autorzy badania w ogóle nie podjęli się przeanalizowania tej próbki o wielkości główki szpilki w poszukiwaniu nowych interakcji między białkami. Niebezpiecznie jest ekstrapolować na „zyliony” rzędów wielkości (z 1020 na 1040) nawet w najlepszej z możliwych sytuacji. Doktor Behe zrobił to jednak w przypadku parametru, który w ogóle nie był przedmiotem zainteresowania. Badacze nie zaprojektowali eksperymentu w celu poszukiwania nowych interakcji między białkami w całym zasobie białek w komórkach – sprawdzali tylko jeden konkretny fenotyp. Behe słusznie zauważył, że nie znaleźli oni takich interakcji, ale wniosek, że nie znaleźli oni ŻADNYCH nowych interakcji między białkami, jest naciągany, jeżeli weźmiemy pod uwagę, iż badacze ich nie szukali. Interesowała ich tylko mała liczba wysoce specyficznych zmian, nie zaś cały proteom. To prawda, że w tym konkretnym przypadku nie odnotowano znalezienia korzystnych mutacji w badanych próbkach, ale wniosek, iż do takich mutacji nigdy nie może dojść albo dochodzi do nich rzadko, to co najwyżej optymistyczna ekstrapolacja.

Oryginał: Is There an Edge to Evolution?, „BioLogos” 2010, December 4 [dostęp: 1 IV 2025].
Przekład z języka angielskiego: Dariusz Sagan
Przypisy
- Por. też części 1–4: D. Ussery, Czy istnieje granica ewolucji? Analiza książki Michaela J. Behego „Granica ewolucji” – rozdział 1, tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 26 marca [dostęp: 3 IV 2025]; tenże, Czy istnieje granica ewolucji? Rozdziały 2 i 3 (analiza), tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 28 marca [dostęp: 3 IV 2025]; tenże, Czy istnieje granica ewolucji? Rozdziały 4 i 5 (analiza), tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 2 kwietnia [dostęp: 3 IV 2025]; tenże, Czy istnieje granica ewolucji? Rozdziały 6 i 7 (analiza), tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 4 kwietnia [dostęp: 3 IV 2025] (przyp. tłum.).
- Por. M.J. Behe, Granica ewolucji. W poszukiwaniu ograniczeń darwinizmu, tłum. Z. Kościuk, „Seria Inteligentny Projekt”, Fundacja En Arche, Warszawa 2020 (przyp. tłum.).
- Tamże, s. 174.
- Tamże, s. 185.
- Por. N.C. Kyrpides, Fifteen Years of Microbial Genomics: Meeting the Challenges and Fulfilling the Dream, „Nature Biotechnology” 2009, Vol. 27, No. 7, s. 627–632, https://doi.org/10.1038/nbt.1552.
- M.J. Behe, Granica ewolucji, s. 179.
- Por. J. Von Eichborn, S. Günther, R. Preissner, Structural Features and Evolution of Protein-Protein Interactions, „Genome Informatics” 2010, January, s. 1–10, https://doi.org/10.1142/9781848165786_0001 (przyp. tłum.).
- Tamże, s. 1 (przyp. tłum.).
- Autor podaje następujący adres internetowy do artykułu Kelsey Luomy na stronie BioLogos: https://biologos.org/articles/a-students-review-of-behes-two-binding-site-rule/. Wygląda jednak na to, że tego artykułu nie ma już na stronie BioLogos, nawet pod zmienionym adresem internetowym (przyp. tłum.).
- M.J. Behe, Granica ewolucji, s. 175.
Literatura:
1. Behe M.J., Granica ewolucji. W poszukiwaniu ograniczeń darwinizmu, tłum. Z. Kościuk, „Seria Inteligentny Projekt”, Fundacja En Arche, Warszawa 2020.
2. Kyrpides N.C., Fifteen Years of Microbial Genomics: Meeting the Challenges and Fulfilling the Dream, „Nature Biotechnology” 2009, Vol. 27, No. 7, s. 627–632, https://doi.org/10.1038/nbt.1552.
3. Ussery D., Czy istnieje granica ewolucji? Analiza książki Michaela J. Behego „Granica ewolucji” – rozdział 1, tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 26 marca [dostęp: 3 IV 2025].
4. Ussery D., Czy istnieje granica ewolucji? Rozdziały 2 i 3 (analiza), tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 28 marca [dostęp: 3 IV 2025].
5. Ussery D., Czy istnieje granica ewolucji? Rozdziały 4 i 5 (analiza), tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 2 kwietnia [dostęp: 3 IV 2025].
6. Ussery D., Czy istnieje granica ewolucji? Rozdziały 6 i 7 (analiza), tłum. D. Sagan, „W Poszukiwaniu Projektu” 2025, 4 kwietnia [dostęp: 3 IV 2025].
7. Von Eichborn J., Günther S., Preissner R., Structural Features and Evolution of Protein-Protein Interactions, „Genome Informatics” 2010, January, s. 1–10, https://doi.org/10.1142/9781848165786_0001.