
Nie ma nic zaskakującego w tym, że przez tysiące lat ludzie byli przekonani, iż Słońce i gwiazdy krążą wokół Ziemi. Widzimy to na własne oczy codziennie. Jednak w 1543 roku Mikołaj Kopernik zaproponował tezę, że to Ziemia krąży wokół Słońca ‒ i w ten sposób zrewolucjonizował nasze rozumienie Układu Słonecznego i Wszechświata. Półtora wieku później Isaak Newton odkrył prawa dynamiki wyjaśniające zachowanie się wszystkich ciał materialnych – nie tylko na Ziemi, ale także w przestrzeni kosmicznej – poprzez odniesienie do działania cząstek materialnych (mas) i sił.
Udoskonalone teleskopy pozwoliły astronomom lepiej poznać inne planety i gwiazdy, dzięki czemu stało się oczywiste, że Ziemia jest tylko znikomo małą częścią Wszechświata. Nie ma w tym nic zaskakującego, że współcześni naukowcy traktują Wszechświat jako najogólniejszy przejaw istnienia praw przyrody, a życie na Ziemi przedstawiane jest jako przypadek szczególny. Większość współczesnych biologów jest zdania, że organizmy żywe są przypadkiem szczególnym (bardzo złożonym) działania praw przyrody opisujących wszystkie zjawiska za pomocą cząstek materialnych i sił.
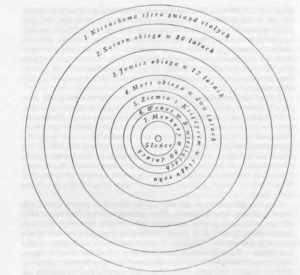
Rycina z: M. Kopernik, O obrotach sfer niebieskich, tłum. M. Brożek, Warszawa – Kraków 1976, s. 22.
Podejście mechanistyczne doskonale odzwierciedla poglądy Kartezjusza, który w XVII wieku zaproponował „metaforę maszyny” – ciała zwierząt zostały przyrównane do maszyn składających się z mniejszych maszyn. Ta metafora ciągle dominuje we współczesnej biologii. W 1998 roku Bruce Alberts, prezydent U.S. National Academy of Sciences [Narodowa Akademia Nauk], napisał w prestiżowym czasopiśmie „Cell”, że „cała komórka [organizmu] może być przyrównana do fabryki z rozbudowaną siecią współpracujących ze sobą linii montażowych, z których każda jest dużym zestawem maszyn białkowych”1. W tym samym wydaniu tego czasopisma znajdują się artykuły o „maszynach modyfikujących chromatyny”2, „maszynach opiekuńczych”3 czy „maszynach wewnątrz maszyn”4. Jeden z artykułów został zatytułowany Mechanical Devices of the Spliceosome: Motors, Clocks, Springs, and Things [Urządzenia mechaniczne spliceosomów: silniki, zegary, sprężyny i części]5.
Darwin i projekt
Nie dość, że badania większości biologów wpisują się w ramy metafory maszyny, to poruszają się oni również w ramach materialistycznego układu odniesienia darwinowskiej teorii ewolucji. Według Darwina wszystkie istoty żywe są potomkami jednego lub kilku wspólnych przodków, którzy ulegli modyfikacjom w wyniku działania niekierowanych procesów, jak losowa zmienność i dobór naturalny. Adaptacje, które dla wcześniejszych biologów były tworem projektu, Darwin tłumaczył doborem naturalnym, czyli procesem działającym bez planu i celu. W 2007 roku Francisco J. Ayala napisał, że Darwin chciał „wyjaśnić projekt organizmów, ich złożoność, różnorodność i pomysłową konstrukcję, jako wynik działania procesów przyrodniczych”, bez udziału inteligencji. Ayala określił to „największym osiągnięciem Darwina”6.
Niemniej Darwin nie wiedział nic o mechanizmach dziedziczenia i powstawania nowych odmian, dlatego też jego teoria miała poważne luki. Na początku XX wieku wydawało się, że genetyka mendlowska wypełni pierwszą lukę, podczas gdy kolejna miała zostać zapełniona w drugiej połowie XX wieku wraz z odkryciem mutacji DNA. W ten sposób powstała tak zwana nowoczesna synteza, w ramach której teorię Darwina połączono z koncepcją, że rozwój organizmów jest sterowany przez program zapisany w sekwencjach DNA. Mutacje DNA mogą zmienić ten program, dostarczając tym samym materiału dla ewolucji. Zdaniem biologa molekularnego Jacquesa Monoda: „dzięki wiedzy o mutacjach i fizycznych podstawach ich losowości, o czym również dowiedzieliśmy się za sprawą biologii molekularnej, mechanizm darwinowski stoi wreszcie na solidnych podstawach. Człowiek musi zrozumieć, że jest tylko przypadkiem”7.
Wielu biologów – prowadzących badania w ramach teorii ewolucji Darwina i biologii molekularnej – traktuje organizmy żywe jako szczególny rodzaj maszyny, coś w rodzaju komputera, w którym sekwencje DNA pełnią funkcję oprogramowania. W podobnym tonie wyraził się Bill Gates, który w 1995 roku oświadczył, że: „DNA jest jak program komputerowy, jednak o wiele bardziej skomplikowany i zaawansowany niż jakiekolwiek oprogramowanie stworzone dotychczas przez człowieka”8. Określenia, takie jak „program genetyczny” [genetic program] i „schemat DNA” [DNA blueprint], przeniknęły zarówno do literatury technicznej, jak i popularnej. Francis S. Collins, kierownik Human Genome Project [Projekt poznania ludzkiego genomu], napisał w 2006 roku w książce Język Boga, że DNA jest „fascynującym tekstem, zawierającym wszystkie instrukcje konieczne do zbudowania ludzkiego ciała”9.
Połączenie pojęć darwinowskiej ewolucji i programu genetycznego prowadzi nas jednak do paradoksu. Komputery i programy komputerowe (tak jak w ogólności maszyny) tworzone są przez istoty inteligentne – ludzi. Nie powinno dziwić, że według zwolenników teorii inteligentnego projektu (ID) organizmy żywe przypominają pod pewnymi względami maszyny i kody, a fakt ten wskazuje na projekt, którego istnienie tak bardzo chcieliby wykluczyć uczeni opowiadający się za ewolucjonizmem darwinowskim. Na przykład Michael J. Behe wskazuje na pewną maszynę molekularną – wić bakteryjną – która może sprawnie funkcjonować tylko wówczas, gdy nie brakuje żadnej z kilkudziesięciu jej części – a to jest cecha charakterystyczna dla inteligentnego projektu10. Stephen C. Meyer zwraca natomiast uwagę na sekwencje DNA cechujące się złożonością i wysokim stopniem wyspecyfikowania, które – analogicznie do programów komputerowych – nie mogły powstać przypadkowo, lecz dają podstawę do wniosku o udziale inteligentnego projektanta11.
Massimo Pigliucci i Maarten Boudry – filozofowie przychylnie nastawieni do teorii ewolucji – napisali, że „kreacjoniści i ich współcześni następcy z Ruchu Inteligentnego Projektu chętnie wykorzystują metaforę maszyny dla własnych celów”12. Dlatego „jeżeli chcemy utrzymać teorię inteligentnego projektu z dala od sal wykładowych, to musimy nie tylko usunąć tę «teorię» z programu nauk biologicznych, ale powinniśmy także rozważnie używać metafor, które mogą powodować wrażenie, że organizmy zostały zaprojektowane”13. Pigliucci i Boudry konkludują, że skoro „metafory maszyny/informacji są wodą na młyn dla kreacjonizmu ID, potęgując intuicje projektu i inne nieporozumienia związane z systemami żywymi, to należy z nich zrezygnować”14.
Istnieją jednak lepsze powody, które skłaniają do odrzucenia metafory maszyny (o niektórych pisali też Pigliucci i Boudry). Chociaż podejście mechanistyczne przyniosło pewne korzyści w badaniach biologicznych, to prawda jest taka, że istoty żywe bardzo różnią się od maszyn.
Koniec z metaforą maszyny
Isaak Newton zaproponował mechanistyczny obraz Wszechświata, który rozbudził wyobraźnię uczonych. Jednak sto lat później filozof Immanuel Kant wskazał, że istoty żywe – nazywał je „istotami organicznymi”15 – nie mogą być ujmowane w kategoriach mechanistycznych. Maszyny organizowane są przez czynnik zewnętrzny, natomiast istoty żywe organizują się same od środka. Kant pisał: „Istota organiczna jest […] nie tylko maszyną, bo ta zawiera w sobie wyłącznie siłę poruszającą, ale posiada jeszcze w sobie siłę kształtującą i to taką, której udziela pozbawionym jej materiom (uorganicznia je)”16. Dlatego też nie sposób właściwie zrozumieć istot żywych poprzez analogię do jakiejkolwiek znanej nam przyczynowości naturalnej. Zamiast tego musimy zdać sobie sprawę, że posiadają one „celowość wewnętrzną”17. Filozof pisał, że wewnątrz organizmu „nic nie jest […] daremne, bezcelowe, nic też nie może być przypisane ślepemu mechanizmowi przyrody”Kant, Krytyka władzy sądzenia, s. 259 (przyp. tłum.).
Kant twierdził, że pogląd o wewnętrznej celowości organizmów jest „ideą regulatywną”18, która rządzi naszym sposobem myślenia. Aby należycie zrozumieć organizmy, trzeba myśleć o nich w kategoriach celu i projektu (współcześni biolodzy, pomimo przyjęcia idei Darwina, mówią w ten sposób o organizmach). Kant nie twierdził jednak, że jego opis odnosi się do „rzeczy samej w sobie”, która jego zdaniem była niepoznawalna – ten pogląd filozoficzny wywarł duży wpływ, chociaż był kontrowersyjny. Najistotniejsze w tym miejscu jest to, że Kant zdawał sobie sprawę, iż organizmy nie mogą być traktowane jako maszyny.
Obecnie biologowie i filozofowie biologii krytykują metaforę maszyny coraz częściej. Keith Baverstock twierdzi, że „szybko rosnący zbiór danych empirycznych podważa dogmatycznie przyjmowane tezy o centralnym znaczeniu DNA, odgrywającym dominującą rolę w procesie ewolucji i regulacji komórkowej – takie ujęcie zależne jest od metafory maszyny”19. Daniel Nicholson napisał, że „pomimo pewnych interesujących podobieństw, organizmy i maszyny to zupełnie różne systemy […], te pierwsze mają celowość wewnętrzną, a drugie – celowość zewnętrzną”20. W związku z tym metafora maszyny „nie stanowi właściwego teoretycznego ujęcia natury systemów żywych”21. Według Ann Gauger „metafora maszyny zawiodła” częściowo dlatego, że organizmy żywe są „istotami, w przypadku których zachodzi przyczynowość o charakterze kolistym” [causally circular beings]22. Do powstania nowych komórek niezbędne są nie tylko istniejące już komórki, ale w wielu przypadkach również szlak biosyntezy danej cząsteczki wymaga udziału takiej samej cząsteczki, jaka jest właśnie syntetyzowana. Stephen Talbott stwierdził otwarcie, że stosowanie metafory maszyny jest „niewybaczalnym błędem”, który „od dziesięcioleci jest powielany przez społeczność uczonych i który wstrzymuje rozwój wiedzy biologicznej”23.
Jednak wyjście poza podejście mechanistyczne w przypadku istot żywych nie będzie łatwe, ponieważ świat mechaniki newtonowskiej wydaje się taki… naturalny. Ale tak jak fizyka relatywistyczna i teoria kwantów wykazały, że mechanika Newtonowska stanowi przypadek szczególny w skalach kosmicznej i atomowej, tak współczesna rewolucja w biologii wskazuje, że mechanika Newtonowska jest szczególnym przypadkiem również w skali organizmów. Wszystkie te trzy rewolucje [relatywistyczna, kwantowa i biologiczna ‒ przyp. tłum.] pokazują, że prawa mechaniki Newtonowskiej stanowią jedynie podzbiór praw rządzących zjawiskami w przyrodzie, która jest bardziej złożona (i trudniejsza do wyjaśnienia), niż zakłada podejście mechanistyczne.
Przewrót w biologii
W latach pięćdziesiątych XX wieku Nicolas Rashevsky prowadził badania w ramach tzw. biologii relacyjnej [relational biology], która miała przezwyciężyć ograniczenia podejścia mechanistycznego w stosunku do istot żywych24. Rashevsky nie analizował składników molekularnych komórki, ale skupił się na wzajemnych relacjach elementów wewnątrz komórki. Jego uczeń Robert Rosen rozwinął biologię relacyjną dzięki wykorzystaniu „teorii kategorii” [category theory]25, stanowiącej podejście matematyczne zaproponowane w latach czterdziestych przez Samuela Eilenberga i Saundersa MacLane’a26. Biologia relacyjna była następnie rozwijana przez Iona Baianu, Andrea Ehresmanna oraz Jeana-Paula Vanbremeerscha27, Paula Kainena28, Aloisiusa Louie29, a także Richarda Sternberga30.
Zdaniem Rosena biologia relacyjna ma charakter rewolucyjny, ponieważ opisuje istoty żywe za pomocą praw, które nie występują w przyrodzie nieożywionej. Przy takim podejściu organizm nie jest już traktowany jako szczególny przypadek działania uniwersalnych praw przyrody, lecz stanowi przypadek ogólny, a przypadkiem szczególnym jest przyroda nieożywiona, ponieważ obowiązuje w niej jedynie podzbiór tych wszystkich praw, które odnoszą się do istot żywych. Rosen wyraził się w sposób następujący:
Organizmy nie są przypadkiem szczególnym ucieleśniającym ogólniejsze zasady czy prawa, które w naszym przekonaniu już znamy. Istoty żywe są świadectwem, że te ogólne prawa są w najwyższym stopniu niekompletne. Obraz Wszechświata opisywanego za pomocą tych praw jest bardzo ubogi i nie obejmuje wszystkich zjawisk przyrodniczych. […] Krótko mówiąc, biologia nie jest szczególnym przypadkiem tych praw i nie jest do nich redukowalna, lecz dostarcza najbardziej spektakularnych przykładów ich nieadekwatności. Alternatywą jest […] ogólniejsze spojrzenie na sam świat nauki, w świetle którego to właśnie prawa mechanistyczne odgrywają rolę przypadków szczególnych31.
Czy to oznacza, że już zrozumieliśmy, czym jest życie? Oczywiście, że nie – tak jak nie zrozumieliśmy Wszechświata po 1543 roku. Ale ta radykalna zmiana perspektywy zaproponowana przez biologię relacyjną – analogicznie jak zmiana perspektywy zaproponowana przez Kopernika – przynajmniej otwiera drzwi do bardziej owocnych poszukiwań.
Czy powyższe ma negatywny wpływ na teorię inteligentnego projektu? W żadnym wypadku.
William A. Dembski w książce Being As Communion: A Metaphysics of Information [Bycie jako relacyjność. Metafizyka informacji] definiuje teorię inteligentnego projektu jako „analizę wzorców (stąd słowo «projekt») w przyrodzie, które świadczą o udziale czynnika teleologicznego (stąd słowo «inteligentny»)”32. Definicja ta nie wymusza jednak utożsamienia tego teleologicznego czynnika z istotą znajdującą się poza obrębem przyrody.
Zdaniem Arystotelesa „projekt” oznaczało „zasadę ruchu w czymś innym niż w rzeczy w ruchu”, podczas gdy „natura” oznaczała „zasadę w samej tej rzeczy”33. Twierdził on, że w obu tych przypadkach „zasada” miała charakter teleologiczny, dlatego też wprowadził rozróżnienie pomiędzy teleologią zewnętrzną i wewnętrzną. Materializm odziera przyrodę z teleologii wewnętrznej, a organizmy traktowane są w jego ramach jak maszyny, ale pozostawia miejsce dla teleologii zewnętrznej. Zdaniem Dembskiego teoria ID nie ogranicza się tylko do teleologii zewnętrznej, ale (jak metafizyka Arystotelesa) obejmuje również teleologię wewnętrzną. Kiedy teoretycy projektu patrzą na życie z perspektywy mechanistycznej, to mają na myśli „środek tymczasowy, który na podstawie argumentu reductio ad absurdum ma doprowadzić do obalenia materializmu. Wówczas zwolennicy teorii inteligentnego projektu będą mogli porzucić mechanistyczne rozumienie życia i dzięki temu uzyskać jego prawdziwy obraz”34.
Dembski twierdzi ponadto, że podstawą życia – jak również całego kosmosu – jest informacja. Definiuje on informację jako realizację określonych możliwości i wykluczenie pozostałych. Ponieważ materia istnieje tylko w postaci obiektów materialnych – czyli jako konkretne realizacje wielu możliwości – więc pod względem ontologicznym informacja wyprzedza materię. Dlatego też „informację należy traktować jako podstawowy byt i przedmiot nauki, a materię trzeba pozbawić jej obecnego, uprzywilejowanego statusu […]. Materialiści uznają, że świat przyrody jest materialny w każdym swoim aspekcie. Zwolennicy realizmu informacyjnego, tacy jak ja, uważają natomiast, że podstawą całego świata przyrody jest informacja”35.
Co jest źródłem informacji w przyrodzie? Zgodnie z darwinowską teorią ewolucji źródłem tym jest dobór naturalny. Jednak Dembski pokazuje, że dobór naturalny jest w gruncie rzeczy „redystrybutorem informacji, a nie jej generatorem czy kreatorem […]. Zgodnie z ujęciem materialistycznym inteligencja nie jest rzeczywista, lecz stanowi epifenomen [zjawisko wtórne ‒ przyp. tłum.] procesów naturalnych. Jeżeli jednak inteligencja jest realna i posiada właściwe sobie zdolności sprawcze, to może robić coś więcej, niż tylko redystrybuować informację – może także ją tworzyć”36. W gruncie rzeczy „definicyjną cechą inteligencji jest zdolność do tworzenia informacji” i „jest ona ostatecznym źródłem informacji”37.
Dembski doszedł do następującego wniosku: „Informacja jest w zasadzie relacyjna, ponieważ powstaje wtedy, gdy niektóre możliwości są realizowane kosztem pozostałych – możliwości związane z informacją istnieją tylko w odniesieniu do innych możliwości”38. Dlatego też „realizm informacyjny […] jest ontologią relacyjną”39. Podobnie jak w przypadku biologii relacyjnej, w myśl realizmu informacyjnego relacje pomiędzy przedmiotami są ważniejsze niż same te przedmioty.
Realizm informacyjny oraz biologia relacyjna, w przeciwieństwie do metafory maszyny i ewolucjonizmu materialistycznego, pozwalają postrzegać organizmy takimi, jakimi one są. Zamiast biologii ewolucyjnej otrzymujemy biologię rewolucyjną.
Jonathan Wells
Oryginał: Revolutionary Biology, „Evolution News & Science Today” 2014, October 20 [dostęp 30 I 2020].
Przekład z języka angielskiego: Robert Olender
Źródło zdjęcia: Pixabay
Ostatnia aktualizacja strony: 30.01.2020
Literatura:
- Alberts B., The Cell As a Collection of Protein Machines: Preparing the Next Generation of Molecular Biologists, „Cell” 1998, Vol. 92, No. 3, s. 291‒294 [dostęp 28 XI 2019].
- Arystoteles, Metafizyka, tłum. K. Leśniak [dostęp 5 XII 2019].
- Ayala F.J., Największe odkrycie Darwina: projekt bez projektanta, tłum. N. Górska i in., „Filozoficzne Aspekty Genezy” 2012, t. 9, s. 7‒32 [dostęp 28 XI 2019].
- Baker T.A., Bell S.P., Polymerases and the Replisome: Machines within Machines, „Cell” 1998, Vol. 92, No. 3, s. 295‒305 [dostęp 28 XI 2019].
- Baverstock, Life As Physics and Chemistry: A System View of Biology, „Progress in Biophysics and Molecular Biology” 2013, Vol. 111, No. 2‒3, s. 108‒115 [dostęp 28 XI 2019].
- Behe M.J., Czarna skrzynka Darwina. Biochemiczne wyzwanie dla ewolucjonizmu, tłum. D. Sagan, „Biblioteka Filozoficznych Aspektów Genezy”, t. 4, Warszawa 2008 [dostęp 28 XI 2019].
- Bukau B., Horwich A.L., The Hsp70 and Hsp60 Chaperone Machines, „Cell” 1998, Vol. 92, No. 3, s. 351‒366 [dostęp 28 XI 2019].
- Collins F.S., Język Boga. Kod życia – nauka potwierdza wiarę, tłum. M. Yamazaki, Warszawa 2008.
- Dembski W., Being As Communion: A Metaphysics of Information, „Routledge Science and Religion Series”, Abingdon – New York 2014 [dostęp 5 XII 2019].
- Ehresmann A.C., Vanbremeersch J.P., Memory Evolutive Systems; Hierarchy, Emergence, Cognition, „Studies in Multidisciplinarity”, Vol. 4, Amsterdam – Oxford 2007 [dostęp 5 XII 2019].
- Eilenberg S., MacLane S., General Theory of Natural Equivalences, „Transactions of the American Mathematical Society” 1945, Vol. 58, No. 2, s. 231‒294 [dostęp 5 XII 2019].
- Gates B., Droga ku przyszłości, tłum. Cichawa, Warszawa 1997.
- Gauger A., Life, Purpose, Mind: Where the Machine Metaphor Fails, „Evolution News & Science Today” 2011, June 1 [dostęp 5 XII 2019].
- Judson H.F., The Eighth Day of Creation: Makers of the Revolution in Biology, New York 1979.
- Kadonaga J.T., Eukaryotic Transcription: An Interlaced Network of Transcription Factors and Chromatin-Modifying Machines, „Cell” 1998, Vol. 92, No. 3, s. 307‒313 [dostęp 28 XI 2019].
- Kainen P.C., On the Ehresmann–Vanbremeersch Theory and Mathematical Biology, „Axiomathes” 2009, Vol. 19, No. 3, s. 225‒244 [dostęp 5 XII 2019].
- Kant I., Krytyka władzy sądzenia, tłum. M. Żelazny, „Dzieła zebrane”, t. IV, Toruń 2014.
- Louie A.H., More Than Life Itself: A Synthetic Continuation in Relational Biology, „Categories”, 1, Frankfurt 2009 [dostęp 5 XII 2019].
- Louie A.H., Relational Biology [dostęp 29 XI 2019].
- Louie A.H., Mathematical Biologist.
- Meyer S.C., Signature in the Cell: DNA and the Evidence for Intelligent Design, New York 2009 [dostęp 28 XI 2019].
- Nicholson D.J., The Machine Conception of the Organism in Development and Evolution: A Critical Analysis, „Studies in History and Philosophy of Biological and Biomedical Sciences” 2014, Vol. 48, s. 162‒174 [dostęp 5 XII 2019].
- Pigliucci M., Boudry M., Why Machine-Information Metaphors are Bad for Science and Science Education, „Science & Education” 2011, Vol. 20, No. 5‒6, s. 453‒471 [dostęp 28 XI 2019].
- Rosen R., Life Itself: A Comprehensive Inquiry Into the Nature, Origin, and Fabrication of Life, „Complexity in Ecological Systems”, New York 1991.
- Staley J.P., Guthrie C., Mechanical Devices of the Spliceosome: Motors, Clocks, Springs, and Things, „Cell” 1998, Vol. 92, No. 3, s. 315‒326 [dostęp 28 XI 2019].
- Sternberg R., DNA Codes and Information: Formal Structures and Relational Causes, „Acta Biotheoretica” 2008, Vol. 56, No. 3, s. 205‒232 [dostęp 5 XII 2019].
- Talbott, Biology’s Shameful Refusal to Disown the Machine-Organism, The Nature Institute, 2014, April 24 [dostęp 5 XII 2019].
Przypisy
- B. Alberts, The Cell As a Collection of Protein Machines: Preparing the Next Generation of Molecular Biologists, „Cell” 1998, Vol. 92, No. 3, s. 291 [291‒294] [dostęp 28 XI 2019].
- Por. J.T. Kadonaga, Eukaryotic Transcription: An Interlaced Network of Transcription Factors and Chromatin-Modifying Machines, „Cell” 1998, Vol. 92, No. 3, s. 307‒313 [dostęp 28 XI 2019] (przyp. tłum.).
- Por. B. Bukau, A.L. Horwich, The Hsp70 and Hsp60 Chaperone Machines, „Cell” 1998, Vol. 92, No. 3, s. 351‒366 [dostęp 28 XI 2019] (przyp. tłum.).
- Por. T.A. Baker, S.P. Bell, Polymerases and the Replisome: Machines within Machines, „Cell” 1998, Vol. 92, No. 3, s. 295‒305 [dostęp 28 XI 2019] (przyp. tłum.).
- Por. J.P. Staley, C. Guthrie, Mechanical Devices of the Spliceosome: Motors, Clocks, Springs, and Things, „Cell” 1998, Vol. 92, No. 3, s. 315‒326 [dostęp 28 XI 2019] (przyp. tłum.).
- F.J. Ayala, Największe odkrycie Darwina: projekt bez projektanta, tłum. N. Górska i in., „Filozoficzne Aspekty Genezy” 2012, t. 9, s. 10 [7‒32] [dostęp 28 XI 2019].
- Wypowiedź Monoda cytowana w: H.F. Judson, The Eighth Day of Creation: Makers of the Revolution in Biology, New York 1979, s. 217.
- B. Gates, Droga ku przyszłości, tłum. P. Cichawa, Warszawa 1997, s. 235.
- F.S. Collins, Język Boga. Kod życia – nauka potwierdza wiarę, tłum. M. Yamazaki, Warszawa 2008, s. 9.
- Por. M.J. Behe, Czarna skrzynka Darwina. Biochemiczne wyzwanie dla ewolucjonizmu, tłum. D. Sagan, „Biblioteka Filozoficznych Aspektów Genezy”, t. 4, Warszawa 2008 [dostęp 28 XI 2019].
- Por. S.C. Meyer, Signature in the Cell: DNA and the Evidence for Intelligent Design, New York 2009 [dostęp 28 XI 2019].
- M. Pigliucci, M. Boudry, Why Machine-Information Metaphors are Bad for Science and Science Education, „Science & Education” 2011, Vol. 20, No. 5‒6, s. 459 [453‒471] [dostęp 28 XI 2019].
- Pigliucci, Boudry, Why Machine-Information, s. 454 (przyp. tłum.).
- Pigliucci, Boudry, Why Machine-Information, s. 468 (przyp. tłum.).
- Prof. Mirosław Żelazny (we wstępie od tłumacza – I. Kant Krytyka władzy sądzenia, tłum. M. Żelazny „Dzieła zebrane”, t. IV, Toruń 2014, s. 14) zwraca uwagę, że „Zarówno niemiecki termin organisch jak i organiesiert są w Krytyce władzy sądzenia określeniami istoty organicznej. Specyfika tego ostatniego terminu polega na wywodzącej się z greki etymologii: istota <organiczna> to dla Kanta istota <zorganizowana>, a dokładnie <organizująca swe istnienie>”. (przyp. tłum.)
- Kant, Krytyka władzy sądzenia, s. 257 (przyp. tłum.).
- Kant, Krytyka władzy sądzenia, s. 259 (przyp. tłum.).
- Kant, Krytyka władzy sądzenia, s. 22 (przyp. tłum.).
- K. Baverstock, Life As Physics and Chemistry: A System View of Biology, „Progress in Biophysics and Molecular Biology” 2013, Vol. 111, No. 2‒3, s. 111 [108‒115] [dostęp 28 XI 2019].
- D.J. Nicholson, The Machine Conception of the Organism in Development and Evolution: A Critical Analysis, „Studies in History and Philosophy of Biological and Biomedical Sciences” 2014, Vol. 48, s. 163 [162‒174] [dostęp 5 XII 2019].
- Nicholson, The Machine Conception, s. 163.
- A. Gauger, Life, Purpose, Mind: Where the Machine Metaphor Fails, „Evolution News & Science Today” 2011, June 1 [dostęp 5 XII 2019].
- S. Talbott, Biology’s Shameful Refusal to Disown the Machine-Organism, The Nature Institute 2014, April 24 [dostęp 5 XII 2019].
- Por. A.H. Louie, Relational Biology, A.H. Louie, Mathematical Biologist [dostęp 29 XI 2019].
- Por. R. Rosen, Life Itself: A Comprehensive Inquiry Into the Nature, Origin, and Fabrication of Life, „Complexity in Ecological Systems”, New York 1991.
- Por. S. Eilenberg, S. MacLane, General Theory of Natural Equivalences, „Transactions of the American Mathematical Society” 1945, Vol. 58, No. 2, s. 231‒294 [dostęp 5 XII 2019].
- Por. A.C. Ehresmann, J.P. Vanbremeersch, Memory Evolutive Systems; Hierarchy, Emergence, Cognition, „Studies in Multidisciplinarity”, Vol. 4, Amsterdam – Oxford 2007 [dostęp 5 XII 2019].
- Por. P.C. Kainen, On the Ehresmann–Vanbremeersch Theory and Mathematical Biology, „Axiomathes” 2009, Vol. 19, No. 3, s. 225‒244 [dostęp 5 XII 2019].
- Por. A.H. Louie, More Than Life Itself: A Synthetic Continuation in Relational Biology, „Categories”, Vol. 1, Frankfurt 2009 [dostęp 5 XII 2019].
- Por. R. Sternberg, DNA Codes and Information: Formal Structures and Relational Causes, „Acta Biotheoretica” 2008, Vol. 56, No. 3, s. 205‒232 [dostęp 5 XII 2019].
- Por. Rosen, Life Itself, s. 33‒34.
- Por. W. Dembski, Being As Communion: A Metaphysics of Information, „Routledge Science and Religion Series”, Abingdon – New York 2014, s. 58 [dostęp 5 XII 2019].
- Arystoteles, Metafizyka, tłum. K. Leśniak [dostęp 5 XII 2019] (przyp. tłum.).
- Dembski, Being As Communion, s. 62.
- Dembski, Being As Communion, s. 91.
- Dembski, Being As Communion, s. 185.
- Dembski, Being As Communion, s. 186‒187.
- Dembski, Being As Communion, s. 29.
- Dembski, Being As Communion, s. 197.